Artículo en PDF
En el año 2011, la OMS cifra en 8.8 millones de casos, la incidencia de tuberculosis en el mundo, de los que cerca de un millón de casos afectan a a niños, con 130000 fallecimientos al año entre la población pediátrica, se registran además, 1.1 millones de fallecimientos entre la población HIV negativa y 35 millones de fallecimientos al año entre la población HIV positiva. Entre la población pediátrica, la tuberculosis figura entre las 10 patologías con mayor mortalidad.
El bacilo de Calmette-Guerin (BCG) es hasta la fecha la única vacuna disponible frente a la tuberculosis. En los países donde la tuberculosis es endémica, los recién nacidos son inmunizados con BCG al nacimiento, con lo que se consiguen tasas aceptables de protección frente a las formas diseminadas de tuberculosis, en especial meningitis tuberculosa. Sin embargo, la protección conferida por la BCG frente a las formas comunes de tuberculosis del adulto es cuestionable, y en la actualidad existe amplio consenso sobre la protección insuficiente de la BCG en las formas pulmonares del adulto. La demostración de un aumento de cepas de tuberculosis resistentes a los quimioterapicos habituales, y el aumento de incidencia en los países desarrollados favorecida por los flujos migratorios y la población VIH, han impulsado en las últimas dos décadas la investigación sobre nuevas vacunas frente a la tuberculosis.
En la actualidad esta aceptado, que la vacunación con BCG no debe ser suspendida en los países con tuberculosis endémica, por ello se han desarrollado muchos esfuerzos en intentar desarrollar cepas de tuberculosis vivas y atenuadas que aumenten la respuesta inmunitaria, algunas de ellas han demostrado mayor protección y seguridad que la BCG. Otras opciones incluyen modificar genéticamente la vacuna BCG, las tres vacunas desarrolladas mediante recombinación genética son (1):
- rBCG30, Foundation Rockville, USA. Se trata de una vacuna similar a la BCG obtenida por recombinación genética que contiene el gen para el Ag85. El primer ensayo clínico en fase I se realizó en USA en 2004, comprobándose mayor inmunogenicidad que con la BCG.
- Mtb72f (Glaxo-Smith-Kline). Se trata de una combinación de dos antígenos inmunogénicos de M. tuberculosis: Mtb39a y Mtb32a con el adyuvante ASO2a o ASOIb. Se inició su ensayo en fase I en USA y Bélgica en el año 2005 en voluntarios adultos con mantoux negativo, comprobándose que es muy inmunógena y que no induce efectos secundarios. A finales de 2007 esta previsto que comience un ensayo clínico en fase II en sudafrica.
- AERAS-402 Foundation Rockville, USA). Esta vacuna contiene un adenovirus serotipo 35, manipulado genéticamente de forma que es incapaz de replicarse y contiene DNA que expresa una proteína de fusión de 3 antígenos de M. tuberculosis: 85A, 85b y TBI0.4.
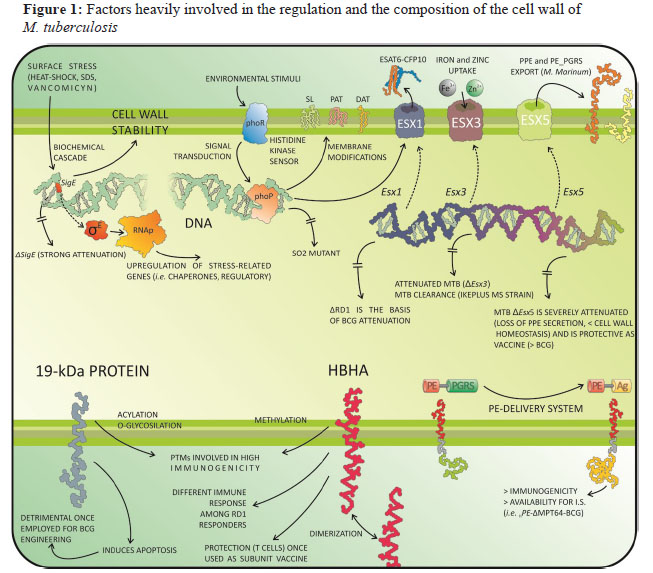
M. tuberculosis es un manipulador activo del sistema inmune del huésped, la explotación de un gran arsenal de biomoléculas le permite interactuar con una amplia gama de ligandos inmunes. La mayoría de estas biomoléculas se localizan en la pared celular de la micobacteria, que tiene una composición muy peculiar y juega un papel fundamental en la patogénesis de las infecciones por micobacterias. Los principales componentes de la pared celular de las micobacterias son lipoarabinomanosidos, arabinogalactano y otros azúcares, ácidos micólicos, glicolípidos y lípidos fenólicos, y peptidoglicano. Esta bien establecido que la pared celular de las micobacterias posee componentes muy inmunogénicos con potentes propiedades inmunoestimulantes, clásicamente resaltados por el uso del adyuvante de Freund, que está hecho de ácido oleico y M. tuberculosis muertos por calor. Las propiedades adyuvantes están vinculadas a la actividad pro-inflamatoria de estas moléculas que inducen TNF, IL-6, IL-1, IL-12, y tienen un efecto regulador del MHC-II y CD1d1 en macrófagos. No es sorprendente que algunos de los antígenos más inmunogénicos de M. tuberculosis se localicen en la pared de la célula o se secreten activamente, mediante sistemas de secreción especializada como el sistema de secreción de tipo siete (T7SS). Los principales componentes de la pared de M. tuberculosis que pueden ser explotadas para diseñar nuevas vacunas vivas atenuadas se muestran en la Figura (2).
Proteína 19-kDa.
La relevancia de las lipoproteínas en la fisiología bacteriana y su potencial para servir como factores de virulencia ha llevado a muchos investigadores a concentrar su atención en éstos componentes bacterianos. Muchas de las lipoproteínas sirven de blanco de la respuesta inmune innata y adquirida, y representan una clase importante de proteínas de la envoltura celular que participa en las interacciones entre el microorganismo y el huésped. Entre estas proteínas, una de las más estudiadas es el antígeno 19-kDa codificado por el gen Rv3763 de M. tuberculosis. La glycolipoproteina 19-kDa se expresa abundantemente en M. tuberculosis, donde se secreta o se encuentra en asociación con la pared celular. Al igual que otros antígenos de micobacterias tales como HBHA, las modificaciones genéticas post-translacionales tienen un gran impacto sobre la respuesta inmunitaria provocada por la proteína de 19-kDa. Originalmente, esta glycolipoproteina suscitó el interés de la comunidad científica, porque se demostró que los anticuerpos murinos la reconocían como un antígeno principal en extractos crudos de M. tuberculosis y, además, se identificó en otras micobacterias patógenas incluyendo Mycobacterium avium, Mycobacterium intracellulare y Mycobacterium leprae, lo que sugería un papel de 19-kDa como un factor de virulencia. Algunos autores han propuesto un papel pleiotrópico de la proteína 19-kDa en la modulación de la respuesta inmune innata, induciendo genes de citoquinas e IFN-? a la baja. En la actualidad, se propone un mecanismo de inductor de apoptosis a través de la vía TLR-2. El hallazgo de que la proteína de 19 kDa está implicada en la inducción de apoptosis llevó a la idea de que podría ser posible mejorar la inmunidad con el uso de vacunas pro-apoptóticas, que podrían generar una respuesta inmune más efectiva. Sin embargo, la expresión de la proteína de 19-kDa en las micobacterias empleadas como vacunas contra la tuberculosis mostró un efecto negativo en la protección contra los bacilos tuberculosos. La inactivación del gen Rv3763 de la BCG, consiguió un efecto similar al observado tras vacunación con BCG estándar. Tomados en conjunto, estos datos sugieren que la proteína 19-kDa tiene un papel fundamental en la patogenicidad de M. tuberculosis, que funciona para engañar al sistema inmune del huésped. Sin embargo, la proteína 19-kDa no se puede considerar un buen candidato para el desarrollo de una nueva vacuna contra la tuberculosis, ya que la respuesta inmune desencadenada va dirigida a disminuir la capacidad del huésped para controlar la infección por M. tuberculosis.
HBHA
La Hemaglutinina de unión a heparina (HBHA) es una proteína de superficie de 21 kDa que se ha implicado en la patogénesis de la tuberculosis. La HBHA media la adherencia de los bacilos a las células epiteliales y macrófagos, pero no está implicada en la difusión de M. tuberculosis en el sitio de infección primaria. Los primeros intentos de explotar este conocimiento por objeto inducir anticuerpos dirigidos contra la HBHA que podrían neutralizar la unión de M. tuberculosis a las células epiteliales, y prevenir la diseminación bacteriana. Curiosamente, la opsonización de BCG con un anticuerpo monoclonal contra HBHA era capaz, en algunos ensayos, de reducir la diseminación bacteriana desde el pulmón hasta el bazo. Varios de estos estudios mostraron que HBHA está expuesto en la superficie de la bacteria y que los anticuerpos específicos podrían neutralizar las propiedades de adhesión de la bacteria. Posteriormente ha quedado establecido que la actividad protectora de una vacuna basada en HBHA se relaciona con la capacidad para provocar una respuesta celular T específica frente a HBHA y secreción de IFN-g y no con la capacidad de inducir anticuerpos. Estos resultados implican que durante la infección por M. tuberculosis, la respuesta inmune dirigida contra HBHA difiere dependiendo de la situación clínica del paciente. Curiosamente, se encontró que la inmunización intranasal de ratones con HBHA adyuvada con la toxina del cólera inducía una respuesta inmune efectiva, tanto humoral como celular. La protección se midió como una reducción de la difusión bacilos desde el pulmón hasta el bazo. Estos resultados destacan la importancia de la administración mucosa de una vacuna basada en HBHA y sugieren que se debe utilizar el modelo animal de difusión de los bacilos desde el sitio de infección primaria, ya que es un paso primordial en la patogénesis de la TB humana.
ESX T7SS.
La identificación de T7SS en micobacterias proporcionó una nueva comprensión de los mecanismos moleculares asociados con la secreción de la proteína y su impacto en la patogénesis de la tuberculosis. Se han identificado cinco sistemas de secreción ESX (ESX1-5) y su caracterización está abriendo nuevas vías de investigación sobre las complejas interacciones huésped-patógeno. ESX-1, está codificada en la región de diferencia 1 (RD1) y su eliminación es responsable de la atenuación de la cepa de la vacuna BCG. ESX-1 codifica una proteína compleja que garantiza la secreción de las proteínas ESAT-6 y CFP-10, que son las dos proteínas altamente inmunogénicas utilizadas en los tests IGRA para diagnosticar la infección latente de TB. La importancia de la función de ESX-1 en virulencia fue también demostrada en la cepa avirulenta H37Ra de M. tuberculosis que contiene una mutación puntual del gen phoP, que anula la secreción de ESAT-6.
La comprensión de la biología del bacilo de la tuberculosis y de los procesos de interacción huésped-patógeno, unido a la posibilidad de diseñar nuevas cepas de BCG y M. tuberculosis, ha abierto nuevas vías para el diseño de vacunas «inteligentes», capaces de provocar la respuesta inmune asociada con la protección y evitar la inducción de la respuesta inmune del huésped asociada con inmunopatología. Se buscan vacunas mas inmunógenas y menos rectógenas que la BCG actual. La pared celular de la micobacteria contiene moléculas altamente inmunogénicas que pueden modular la respuesta inmune tras la inmunización (2).
Dr. José Uberos
Servicio de Pediatría, Hospital Clínico San Cecilio, Granada
Referencias
(1) Hussey G, Hawkridge T, Hanekom W. Childhood tuberculosis: old and new vaccines. Paediatr Respir Rev 2007 Jun;8(2):148-54.
(2) Morandi M, Sali M, Manganelli R, Delogu G. Exploiting the mycobacterial cell wall to design improved vaccines against tuberculosis. J Infect Dev Ctries 2013;7(3):169-81.